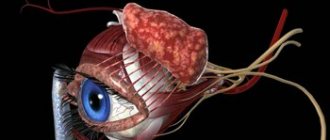
Histology of the retina and its structure
The retina (retina) of the eye
- the inner lining of the eyeball - consists of two parts. The back is photosensitive; the anterior part, which is not sensitive to light, forms the inner lining of the ciliary body and the back of the iris. The retina arises from a protrusion of the anterior medullary vesicle, or forebrain. When such an eye vesicle begins to contact the surface of the ectoderm, it gradually invaginates in the central region, resulting in the formation of a double-walled eye cup.
Outer wall
gives rise to a thin layer which in adults is known as the pigment epithelium; the visual, or functionally active part of the retina - the neural retina - develops from the inner layer.
Pigment epithelium
consists of columnar cells, the nucleus of which is displaced to their basal part. The basal parts of these cells are tightly adjacent to Bruch's membrane, and the cell membranes form numerous basal invaginations. Mitochondria predominate in the cytoplasmic zone near these invaginations. These signs indicate the participation of this zone of cells in ion transport.
On lateral cell membranes
there are intercellular connections with well-developed closure bands and adhesion bands near the apices;
Desmosomes and gap junctions are also identified. These morphological characteristics indicate that the epithelial layer is tightly overlapped in the apical and basal regions and that there is communication between the cells. These specialized connections provide the electrical potential difference that occurs due to ionic transport between the two surfaces of this epithelium. On the apical part of the cells
there are outgrowths of two types: thin and long microvilli and cylindrical layers that cover the ends of the photoreceptors.
Since none of the specified types of outgrowths
not anatomically connected to photoreceptors, these areas can be separated from each other, for example, when retinal detachment develops. This common and serious human condition can be effectively treated using laser surgery.
Cytoplasm of pigment epithelial
contains an extensive aEPS, in which vitamin A is believed to be esterified and transported to photoreceptors.
The apical cytoplasm and microvilli contain numerous melanin granules. Melanin is synthesized in these cells through a mechanism similar to that described in skin melanocytes. The function of this dark pigment is to absorb light after stimulation of photoreceptors. The apical part of the cell
contains numerous dense vesicles of variable shape, which represent various stages of phagocytosis and digestion of the ends of the outer segments of the photoreceptors.
Retina. The cross-section shows most of its components. Color: pararosaniline-toluidi new blue. Low magnification.
Three layers of retinal neurons. The arrows indicate the direction of the incident light. Stimulation of rod and cone cells due to incident light propagates in the opposite direction.
Close connection of Müller cells with neural elements in the photosensitive retina.
Müller cells (dark fibrous cells) are considered the structural and functional equivalent of astrocytes in the central nervous system because they surround and support the neurons and nerve processes of the retina. Visual part of the retina
- its posterior, or light-sensitive, section is a complex structure containing at least 15 types of neurons connected to each other by synapses, which belong to at least 38 different types. The visual part of the retina consists of an outer layer that forms light-sensitive rod and cone cells, an intermediate layer that includes bipolar neurons that connect rod, cone cells and ganglion cells, and an inner layer of ganglion cells that form connections with bipolar cells with their dendrites and send axons into the brain. These axons converge at the optic papilla to form the optic nerve.
Histology.RU
Material taken from the site www.hystology.ru
The wall of the eyeball consists of three membranes - outer, middle and inner. The eyeball also contains light-refracting formations and media - the lens, the fluid of the anterior and posterior chambers of the eye, the vitreous body (Fig. 184).
The outer shell has two parts - the cornea and the sclera.
The cornea (cornea) is the anterior part of the outer shell, consists of the anterior corneal epithelium, basement membrane, anterior limiting membrane, corneal substance proper, posterior limiting membrane and posterior corneal epithelium (Fig. 185).
Multilayered squamous non-keratinizing anterior epithelium (epithelium anterius) consists of 5 - 7 layers of cells. It contains numerous receptor endings that give the cornea greater tactile sensitivity (corneal reflex). Basal epithelial cells have pronounced mitotic activity, so when damaged, the corneal epithelium quickly recovers. The anterior epithelium continues into the epithelium of the conjunctiva and is moistened by the secretion of the lacrimal and conjunctival glands.
The basement membrane is a homogeneous protein-polysaccharide plate.
The anterior limiting membrane (Bowman's) has different thicknesses in different animals; especially pronounced in the cornea of cattle. Electron microscopy revealed thin collagen fibrils in the composition of this membrane, intertwined like felt.
The cornea's own substance (substantia propria corneae) is the bulk of the cornea. It consists of numerous, regularly alternating connective tissue plates, each of which has parallel bundles of collagen fibrils. Between the plates lie flattened cells of the fibroblastic series and contain an amorphous component. The transparency of its own substance is facilitated by the fact that all its collagen fibrils are parallel oriented and equal in thickness, due to which the light they scatter is extinguished as a result of interference. This is also facilitated by sulfated glycosaminoglycans, which determine the tissue’s ability to swell and maintain the ordered arrangement of fibrils. The place of transition of the cornea's own substance into the connective tissue of the sclera is called the limbus (limbus - edge). In the region of the limbus there is also the edge of the anterior limiting membrane, that is, it does not pass from the cornea to the sclera.
The posterior limiting membrane (Descemet's) is a homogeneous plate microscopically. Electron microscopy revealed collagen fibrils in it, which, intersecting, form hexagonal figures.
The squamous epithelium of the posterior surface of the cornea consists of a single layer of cells with hexagonal and other shapes. This epithelium becomes the epithelium covering the anterior surface of the iris.
The cornea provides a significant percentage of focusing
Rice. 184. Diagram of the structure of the eyeball:
1 - edge of the upper eyelid with eyelash; 2 - conjunctiva; 3 - ciliary body and ciliary part of the retina; 4 - sclera (tunica albuginea); 5 - choroid; 6 - retina; 7 - pigment layer; 8 - optic nerve and its membranes; 9 - optic nerve papilla; 10 - vitreous body; 11 - lens; 12 - lens ligament; 13 - anterior and posterior chambers of the eye; 14 - iris; 15 - pupil; 16 - cornea.
Rice. 183. Cornea of a calf eye:
1 - stratified squamous epithelium; 2 - anterior limiting membrane; 3 - intrinsic substance of the cornea; 4 - posterior limiting membrane; 5 - layered squamous epithelium.
eye abilities and acts like a strong magnifying glass. This is a special part of the eye that does not have blood capillaries; it receives nutrition from the anterior chamber of the eye and the vessels of the limbus. Thus, the cornea is sufficiently isolated and it is thanks to this circumstance that it can be transplanted from one organism to another.
In case of damage to the cornea and an inflammatory process, blood capillaries grow into it and cells (leukocytes, etc.) penetrate, which causes a violation of the optical properties and clouding of the cornea.
Sclera (sclera - hard) - opaque posterior and anterolateral parts of the outer shell, white, the strongest in the wall of the eyeball. It consists of dense connective tissue in which collagen fibers and plates formed from them are located parallel to the surface of the eye. Between them are elastic fibers and flattened fibroblasts.
At the border with the cornea, a thickening of the sclera is formed, clearly visible in the form of a roller in carnivores. In the scleral tissue behind the ridge there are small branched cavities - the venous plexus, which ensures the outflow of fluid from the anterior chamber of the eye. At the back of the sclera there is a cribriform plate (lamina cribrosa), containing small openings through which nerve fibers pass, forming the optic nerve. The outside of the sclera is covered with episcleral loose connective tissue containing numerous capillaries.
The sclera serves as a strong skeleton of the eye wall; fibers of the tendons of the eye muscles penetrate it and then attach to it.
In birds, due to incomplete ossification of the orbital walls, the sclera near the cornea contains tiled-like small bone scales, which, when connected, form a kind of protective ring. Distal from it to the point where the optic nerve enters the sclera there is hyaline cartilaginous tissue.
The middle shell consists of three parts: the iris, the ciliary body and the choroid (Fig. 186).
The iris is the anterior part of the middle membrane . The space between the iris and cornea is called the anterior chamber of the eye, and between the iris and the lens is called the posterior chamber. In the central part of the iris there is a hole - the pupil, which is round in shape in dogs, pigs and birds, in the form of a vertical slit in cats, and transversely oval in herbivores. The posterior edge of the iris, connecting it to the ciliary body, is called the ciliary margin. The basis of the iris is made up of bundles of smooth muscle tissue cells and loose connective tissue with a large number of pigment cells -
Rice. 186. Scheme of the structure of the anterior part of the eyeball:
1 - cornea; 2 - anterior chamber of the eye; 3 - iris; 4 - posterior chamber of the eye; 5 - lens; 6 - ciliary band - lens ligament; 7 - vitreous body; 8 - venous sinus of the sclera; 9 - ciliary muscle; 10 - ciliary corolla; 11 - sclera; 12 - jagged line; 13 - retina (Fig. Kozlov).
chromatophores and numerous blood vessels. On a cross-section of the iris in the direction from the anterior to the posterior surface, five layers are distinguished: epithelial, external border, vascular, internal border and pigment. The latter is a continuation of the pigment epithelium of the ciliary body and then the retina. In all layers of the iris, there are pigment cells in varying numbers, which determine the color of the eyes. Albino animals do not have pigment cells, so their iris is red, due to the fact that blood vessels are visible through its thickness.
Smooth muscle tissue forms two muscles in the iris. The muscle that constricts the pupil (sphincter) consists of bundles of cells oriented circularly and located near the pupillary edge of the membrane. The bundles of cells that dilate the pupil (dilator) have a radial direction and are located in the posterior, ciliary zone of the iris. With the help of muscles, the flow of light rays into the eyeball is regulated, that is, the iris acts as a diaphragm. The dilator pupillary muscle is innervated by postganglionic sympathetic fibers of the cranial cervical ganglion, and the pupillary sphincter is innervated by postganglionic parasympathetic fibers of the ciliary ganglion.
In horses and ruminants, at the free pupillary edge of the iris there are outgrowths (grains) penetrated by blood vessels and containing highly pigmented cells.
The ciliary (ciliary) body (corpus ciliare) is a thickened part of the middle membrane, located between the iris and choroid. There is a posterior, thinner part with small folds - the ciliary ring and an anterior, thicker part with high processes directed towards the lens - the ciliary corolla (ciliary crown). The processes and folds of the ciliary body are covered with the ciliary part of the retina - epithelium, which has two layers: an outer layer of pigmented cells and an inner layer of pigmentless cells facing the eye cavity. Epithelial cells take part in the formation of fluid that fills the anterior and posterior chambers of the eye. The bulk of the ciliary body consists of ciliary muscle, formed by bundles of smooth muscle cells located in three directions: annular, radial and meridional. Between the muscle bundles there is connective tissue containing blood capillaries and pigment cells.
Due to the motor activity of the muscles, the ciliary body is of great importance in the accommodation of the eye. When the muscles contract, the tension in the ligament that supports the lens is weakened, and it becomes more rounded, which adapts the eye to viewing objects at close range. When muscles relax, the opposite effect is achieved.
The choroid (tunica vasculosa) is the posterior part of the middle tunica, characterized by an abundance of blood vessels. It consists of connective tissue, in which a network of elastic fibers and many pigment cells are developed. According to the structure of the shell, four plates are distinguished: supravascular, vascular, choriocapillaris and basal. With the help of the supravascular plate, the choroid is connected to the sclera; the vascular plate contains a network of large vessels, and the choriocapllary plate contains a dense network of blood capillaries. Between the vascular and choriocapillary plates in animals there is an avascular zone, consisting of multifaceted cells in carnivores (tapetum lucidum) or interwoven connective tissue fibers (tapetum fibrosum). In cats, the cytoplasm of the tapetum cells contains regularly arranged needle-shaped crystals. It is believed that the presence of this layer is responsible for the glow of the eyes in the dark by reflected light. The basal lamina separates the choroid from the retinal pigment epithelium.
The retina is the inner layer of the wall of the eyeball, adjacent to the vitreous body. In accordance with the location, structure and function, two parts are distinguished in the retina: the neurocellular visual (pars nervosa), lining the back, most of the wall of the eyeball, and the anterior pigment (pars pigmentosa), covering the inside of the ciliary body and the iris. In the visual part the perception of light occurs
Rice. 187. Scheme of the structure of the retina:
A - diagram of the arrangement of neurons in the retina (according to Dowling and Boycott); P - rod cell; K - cone cell; BC - bipolar cell; Horus - horizontal cell; Am - amacrine cell; G - ganglion cells; B — retina on a histological specimen; 1 - layer of pigment epithelial cells; 2 - layer of rods and cones; 3 - external limiting membrane; 4 - outer nuclear layer; 5 - outer mesh layer; 6 - inner nuclear layer; 7 - inner mesh layer; 8 - ganglion layer; 9 - layer of nerve fibers; 10 - internal limiting membrane.
irritations and converting them into a nerve signal. These parts are delimited by a line called the jagged edge.
The visual part consists of two layers: the inner one is photosensitive, containing photoreceptor, primary sensory nerve cells of two types with their complex processes called rods and cones, and the outer one is pigmented.
The light-sensitive layer of the retina contains several types of nerve cells and one type of glial fiber-like cells. The nuclear-containing areas of all cells form three nuclear layers, and the zones of synaptic contacts of cells form two mesh layers. Thus, in the visual part of the retina, when examining its cross section in a light microscope, the following layers are distinguished, counting from the surface in contact with the choroid: a layer of pigment epithelial cells, a layer of rods and cones, an external limiting membrane, an outer nuclear layer, an outer reticular layer, inner nuclear layer, inner reticular layer, ganglion layer, nerve fiber layer and internal limiting membrane (Fig. 187).
The pigment epithelium is the outermost layer of the retina, the cells of which are located at their bases on the basement membrane adjacent to the choroid, and processes extend from the apical surface, located between the outer segments (rods and cones) of light-sensitive cells. The processes of pigment cells contain the pigment melanin, which can move in the cytoplasm and therefore, depending on the lighting, be either in the basal part or in the processes of the cells, absorbing a large (up to 80%) part of the light. In addition, pigment epithelial cells provide nutrients and vitamin A from the choroid to the nerve cells of the retina.
The layer of rods and cones consists of the outer segments of visual (photoreceptor) cells, which are surrounded by processes of pigment cells and are located in a matrix containing glycosaminoglycans and glycoproteins. There are two types of photoreceptor cells, differing not only in the shape of the outer segment, but also in number, distribution in the retina, ultrastructural organization, as well as in the form of synaptic connection with the processes of the deeper located retinal elements following the visual cells - bipolar and horizontal cells. Rods have a higher sensitivity to light and are receptor cells for black-and-white twilight vision, cones - color daytime vision. The retina of diurnal animals and birds (daytime rodents, chickens, pigeons) contains almost exclusively cones; in the retina of nocturnal birds (owls, etc.) visual cells are represented predominantly by rods. Significantly more rods are located on the periphery of the visual part of the retina, which is involved in the visual process in low light.
Each photoreceptor cell consists of outer and inner segments; the rod has a thin, long, cylindrical outer segment, while the cone has a short, conical outer segment. However, it is not always possible to distinguish these cells by the shape of the outer segment. Thus, the cones of the fovea - the place of best perception of visual stimuli - have an elongated thin outer segment and resemble a rod. The internal segments of rods and cones also differ in shape and size; at the cone it is much thicker. The main cellular organelles are concentrated in the internal segment: a cluster of mitochondria, polysomes, elements of the endoplasmic reticulum, and the Golgi complex. In the inner segment of the cones there is a section consisting of a cluster of mitochondria tightly adjacent to each other with a lipid droplet located in the center of this cluster - an ellipsoid (Fig. 188).
Both segments are connected by a so-called stalk, the ultrastructural organization of which is typical for cilia: it contains nine pairs of fibrils located around the circumference, with no neutral pair (9 × 2 + 0). Cilia in optic cells develop from basal bodies located in the epical parts of the inner segment. From basal
Rice. 188.
Scheme of the ultramicroscopic structure of rod-bearing (A) and cone-bearing (B) receptor cells of the retina: 1 - outer segment; 2 - internal segment; 3—nucleus-containing zone; 4 - synaptic zone; a - cilia; b - mitochondria; c — lipid drop; d — endoplasmic reticulum; d - core; e - disks.
The body has a root that extends deep into the inner segment. The outer segments of rod and cone cells contain many disks consisting of double membranes. During embryogenesis, rod and cone discs form as folds of the outer plasma membrane. Then, in the rods, the connection between the discs and the outer membrane is lost, with the exception of a few basal ones. It has been shown that in the formed rod there is a constant formation of new discs by invagination of the plasmalemma in the basal part of the outer segment and subsequent detachment of this invagination.
In cones, the connection between the disc and plasma membranes is maintained along the entire length. The cones of the fovea, as well as the apical portions of the outer segments of primate cones, contain discs detached from the outer membrane. Numerous disks in the volume of the segment are oriented perpendicular to the long axis of the cell. The distance between the disks is the same and is about 30 im. The number of discs in a segment varies greatly in different vertebrates: the outer segment of a frog's rod contains about 1000 - 1,500 discs, and the bull's rod segment contains 200 discs. The discs that fill the outer segments of the rods and cones contain molecules of visual pigments. Of the photopigments, the best studied is rhodopsin, which is found in the rods of all vertebrates. It consists of opsin protein and vitamin A aldehyde - retinal. With a lack of vitamin A, visual perception is impaired, and rod perception is faster than cone perception. There is a particularly high density of rhodopsin molecules in the disc membranes on the side facing the incident light. The absorption of light by the pigment is the first link in the chain of transformations leading to the disintegration and bleaching of the visual pigment, which, in turn, leads to a change in the ionic permeability of the photoreceptor membrane and the appearance of an early receptor potential, that is, the appearance of a visual signal.
The membranes of cone discs contain pigments of different chemical composition. There are three different types of cones, each type containing predominantly only one pigment. The most studied cone pigment is iodopsin. The different visible colors depend on the ratio of the three types of cones stimulated. Color blindness (color blindness) is caused by the absence of one or more types of cones.
Nucleated areas of photoreceptor cells form the outer nuclear layer. The nuclei of cone cells are lighter and larger in comparison with the nuclei of rod cells. The central process of photoreceptor cells in the outer reticular layer comes into contact with the dendrites of bipolar cells and the processes of horizontal cells. Bipolar nerve cells are the next neurons to which impulses are transmitted from light-sensitive cells. With their nuclear-containing areas, bipolar neurocytes form the inner nuclear layer, and the central processes of rod- and cone-shaped cells form synapses with their dendrites. In this case, some bipolar neurocytes contact many cone cells (flat bipolars), others only with one, and still others are associated with rod photoreceptors (rod bipolars).
The nuclear-containing regions of some bipolars are located especially close to the next inner reticular layer. It is believed that these are cells that conduct impulses in the opposite direction - to the visual cells, and are called centrifugal bipolar cells.
In the outer zone of the same inner nuclear layer there are horizontal neurocytes. Their numerous short dendrites are directed towards the light-sensitive cells, and the long axon extends in a horizontal direction and also comes into contact with the central processes of the light-sensitive cells. In the inner zone of the same nuclear layer are located the bodies of another type of cell - amacrine neurocytes. Their highly branching processes in the inner reticular layer form associative connections with the dendrites of ganglion cells. It is believed that horizontal and amacrine neurocytes cause a presynaptic inhibitory effect.
Axons of bipolar cells in the inner reticular layer are involved in the formation of synaptic contacts with the dendrites of ganglion cells. The nuclear-containing areas of which form the ganglion layer of the retina. Ganglion cells are the largest cells; basophilic granularity is well expressed in their cytoplasm. The radially directed axons of the ganglion cells pass through the layer of nerve fibers, become covered with myelin sheaths and converge to the exit site of the optic nerve and form it.
Thus, a chain of three neurons is formed in the retina: photoreceptor (rod and cone cells), bipolar and ganglion. These radially directed chains include horizontal and amacrine cells that form connections in the horizontal direction.
Among neuroglial cells, the most characteristic are fiber-like supporting radial gliocytes (gliocytus radialis). These long and narrow cells extend through the entire thickness of the inner layer perpendicular to the surface of the retina, and the nucleated areas are located in the inner nuclear layer. The outer ends of ray gliocytes form the outer limiting membrane, located between the layer of rods and cones and the outer nuclear layer, and the expanded and tightly adjacent inner ends form the inner limiting membrane, separating the retina from the vitreous body. Along with ray gliocytes, astrocytes and microglial cells are found in the retina.
The arrangement of cells and the thickness of the retina in different parts of its visual part are not the same. In the area of the visual axis projection, the rounded part of the retina is called the macula macula, and the deep central part of the macula macula is called the fovea. At this point, all layers of the retina, with the exception of the outer nuclear layer, are thinned, and the photoreceptor cells are very densely packed cone-bearing cells (there are no rod-bearing cells in the fovea). For this reason, the fovea region gives the best perception of colors and details of objects. However, it is less sensitive to light than the peripheral retina, which has a higher concentration of rod cells. At the point where the fibers that form the optic nerve converge and the blood vessels enter, there is an elevation on the retina. This area, located towards the inner edge of the eye from the macula, is called the blind spot; it has no light-sensitive cells.
Vision analyzer. The nerve signal originating in light-sensitive cells is transmitted to bipolar and from them to ganglion neurocytes, the axons of which form the optic nerve. On the ventral surface of the brain, the optic nerve of the right and left eyes intersect and after the intersection continue in the form of visual pathways to the subcortical centers - the geniculate body of the visual thalamus and the nuclei of the nasal quadrigeminal region. Fibers with axons of cells of the external geniculate body go to the occipital region of the cerebral cortex, which is the cortical center of the visual analyzer. The axons of neurons in the visual cortex form numerous centrifugal pathways. Part of the fibers
Rice. 189. Diagram of the structure of the lens:
1 - capsule; 2 - epithelial cells of the anterior surface; 3 - elongating epithelial cells; 4 - peripheral fibers; 5 - central fibers.
reaches the retina and provides cortical control of the activity of retinal neurons. From the nasal colliculi of the quadrigeminal, fibers form centrifugal pathways along which the impulse is transmitted to the motor cells of the cervicothoracic part of the spinal cord. Through them, reflex movements of the head, neck and eye muscles are carried out. With the participation of neurons of the parasympathetic nucleus (Yakubovich) and neurons of the ciliary ganglion, reflex contractions of the sphincter of the pupil and the muscles of the ciliary body occur.
The light-refracting apparatus of the eye is represented by the cornea, the fluid of the anterior and posterior chambers of the eye, the lens and the vitreous body.
Lens _ A transparent, biconvex lens-shaped formation located between the iris and the vitreous body. Consists of a capsule, epithelial cells and derivatives of these cells, called lens fibers (Fig. 189).
The lens capsule is a homogeneous elastic membrane that surrounds it on all sides. Contains proteins (collagen, glycoproteins) and sulfated glycosaminoglycans. Fibers of the ciliary band of the ligament of cinnamon, coming from the ciliary body, are attached to the outer surface of the capsule along the equator of the lens. When the tension of these fibers is weakened (at the moment of contraction of the ciliary muscle), the lens takes on a more convex shape, which adapts the eye to seeing nearby objects. On the anterior surface under the capsule there is a single-layer cubic epithelium, the cells of which, moving towards the equator, divide, become more elongated, take on a meridional arrangement and behind the equator turn into lens fibers. There are transitional fibers with nuclei and central fibers without nuclei. Each fiber has the appearance of a transparent hexagonal prism; the main chemical substances of their cytoplasm are crystallin proteins.
With age, the lens becomes less elastic, which affects its focusing properties.
The vitreous body (corpus vitreum) is a transparent jelly-like mass that fills the cavity bounded in front by the lens, on the sides by the posterior side of the ligament of zinn, and behind by the internal limiting membrane of the retina. The vitreous body, being one of the main light-refracting media, is also important in maintaining intraocular pressure and ensuring metabolic processes.
From the papilla of the optic nerve of the retina towards the posterior surface of the lens, the hyaloid canal passes through the vitreous body - a remnant of the embryonic vessel of the eye. In birds (geese) there is a special formation in this place - a comb, the front end of which is connected to the lens capsule. It consists of connective tissue and contains blood capillaries. The colloidal mass of the vitreous body contains a complex protein - vitrein and hyaluronic acid. Electron microscopy reveals thin collagen fibers in this mass.
Vascularization . Blood flows to the wall of the eyeball through the central retinal artery and ciliary arteries. The central retinal artery runs inside the optic nerve and splits into capillaries that supply the deep layers of the retina. In the outer nuclear layer and the layer of rods and cones: there are no vessels; these layers receive nutrients from the capillaries of the choroidal plate of the choroid. Blood. from the capillary network it collects into small venous trunks that flow into the central retinal vein.
The ciliary arteries - branches of the ophthalmic artery and arteries of the eye muscles - branch into short and long arteries that supply mainly the choroid, sclera and peripheral parts of the cornea. Blood from the capillaries originating from these arteries is collected in veins that run parallel to the arteries. Their largest branches on the surface of the eyeball are called vortex veins. There are lymphatic spaces in the wall of the eyeball.
Reviews (0)
Add a review
Histology of retinal rod cells and their structure
Rod cells
- thin elongated cells consisting of two sections. The outer photosensitive rod-shaped part consists mainly of numerous (600-1000) flattened membrane disks, assembled like a coin column.
Discs in rod cells
not associated with the plasma membrane; the outer segment and the inner segment are separated from each other by a narrowed portion. Directly below this constriction is the basal body, from which the cilium extends and extends into the outer segment.
Internal segment
rich in glycogen and contains a large accumulation of mitochondria, most of which lie near the constriction. This local accumulation of mitochondria provides the energy needed for vision and protein synthesis. Polyribosomes, present in large quantities below the portion of the inner segment containing mitochondria, are involved in protein synthesis.
Some of these proteins move to the outer segment of rod cells
, where they are incorporated into membrane disks. The flattened disks of rod cells contain visual purple pigment, or rhodopsin, which becomes bleached in light and provides visual stimulation. This substance has a globular molecule and is located on the outer surface of the lipid bilayer of flattened membrane disks.
The human retina contains approximately 120 million rod cells
. They are extremely sensitive to light and are considered to be receptors that are used in low light conditions, such as at dusk or at night. The outer segment is the photosensitive region, and the inner segment contains structures involved in metabolic reactions necessary for the biosynthetic processes and energy production in these cells.
As shown by autoradiographic studies
, proteins found in the vesicles of rod cells are synthesized in the internal segments of these cells, which contain multiple polyribosomes. From here they are transported to the outer segment and accumulate at its basal region, where they are incorporated into membranes containing a bilayer of phospholipids that form flattened discs.
These structures gradually migrate to the cell apex, where they are removed and further phagocytosed and digested by pigment epithelial
. It is estimated that in a monkey, approximately 90 vesicles are formed in each cell per day. The entire migration process, from assembly in the basal region of the cell to removal in the apical region, takes from 9 to 13 days.
Ultrastructure of rod (right) and cone (left) cells. The highlighted rectangular area is shown in the electron micrograph in the figure below.
Retina. At the top are the internal segments. This photosensitive region consists of parallel flat membrane disks. A cluster of mitochondria is located in the inner segment. In the middle there is a basal body with a cilium extending from it, which further changes, turning into an outer segment. Electron micrograph.
The boundary between the photosensitive layer and the retinal pigment epithelium layer. At the bottom of the micrograph are parts of two pigment epithelial cells with specialized connections (C) between their lateral plasmalemmas. Above the pigment cells are the ends of several outer segments of the rod cells, which interdigitate with the apical processes of the pigment epithelial cells (P). Large vacuoles containing flattened membranes (arrows) separated from the ends of the rod cells. L - lysosomal vesicles. Electron micrograph.
Functions of retinal pigment epithelial cells. Note that numerous processes extend from the apical part of the cell, which fill the spaces between the outer segments of the light-sensitive cells, and the membrane of the basal region forms invaginations into the cytoplasm. This type of cell performs several functions, including the synthesis of melanin granules that absorb reflected light in the eye chamber. This process is represented on the right side of the figure, which shows the organelles involved in melanin synthesis. On the left side of the figure, lysosomes containing enzymes synthesized in the granular endoplasmic reticulum (gRES) fuse with and digest the phagocytosed apical sites of photoreceptors. In addition to these functions, pigment cells are likely involved in ion transport as they maintain the electrical potential between the two surfaces of the epithelial membrane. The relatively well-developed agranular endoplasmic reticulum (aER) is involved in the esterification of vitamin A.
Treatment of retinoschisis of the retina
In some cases, the process of retinoschisis stabilizes for a long time or progresses slowly, without significantly affecting the quality of vision. Such patients are required to undergo regular observation by an ophthalmologist; it is possible that a special regimen, supportive drug therapy (absorbable and/or angioprotective drugs, vitamins) or preventive measures will be prescribed.
In other cases, the process rapidly spreads to central areas, creating a real risk of blindness. The main method of treating such developments of retinoschisis is laser coagulation - “fusion” of tissues using a laser (in this case, the procedure is designed to restore the multilayer integrity of the retina). Such intervention is especially indicated in situations where retinoschisis develops against the background of a tendency to retinal detachment (or, conversely, is complicated by detachment). In the most severe cases, when all other measures taken are ineffective, they resort to the surgical operation of vitrectomy - removal of any volume (sometimes complete) of the vitreous body, replacing it with a biocompatible solution and mechanically securing the retina.
As can be seen from the above, retinoschisis is a complex and severe, prognostically unfavorable eye disease, which can be caused by various factors, but in any case should not be left without the control of specialists. Timely seeking qualified ophthalmological care and following all doctor’s instructions significantly increases the chances of maintaining vision, at least at a satisfactory level.
Histology of retinal cone cells and their structure
Cone cells
- just like rod neurons, they are elongated neurons. There are about 6 million cone cells in the human retina. They are similar in structure to rod cells; they also contain outer and inner segments, a basal body with a cilium, and an accumulation of mitochondria and polyribosomes.
Cone cells
differ from rods in the shape (conical) and structure of their outer segments. As in rod cells, this region consists of stacked membrane disks, which, however, are not separated from the outer plasma membrane, but are formed as its invaginations. In cone cells, newly synthesized protein is not concentrated in newly formed discs, as is the case in rod cells, but is distributed evenly throughout the outer segment.
Other retinal cells
Bipolar cell layer
includes two types of cells: diffuse bipolar cells, which form synapses with two or more photoreceptors, and monosynaptic bipolar cells, which establish contact with the axon of only one cone photoreceptor and only one ganglion cell. A certain number of cone cells thus transmit their impulses directly to the brain.
Cells of the ganglion layer
form contacts with bipolar cells; in addition, they send their axons to a special part of the retina, where they unite to form the optic nerve. This receptor-free area is known as the retinal blind spot, optic papilla, or optic nerve head. Ganglion cells are typical nerve cells containing a large euchromatic nucleus and basophilic Nissl bodies.
These cells
, like bipolar cells, based on their connections with other cells, are divided into diffuse and monosynaptic. In addition to these three main cell types (photoreceptor, bipolar and ganglion), there are other cell types that are more diffusely distributed throughout the retinal layers.
Horizontal cells
form contacts between different photoreceptors. Their exact function is unknown, but they may be involved in signal integration.
Amacrine cells
- various types of neurons that provide contacts between ganglion cells. Their function is also unclear.
Supporting cells
belong to neuroglia, which includes, in addition to astrocytes and microglial cells, individual highly branched cells (Müller cells). The processes of these cells connect the neural cells of the retina and extend from the inner to the outer limiting membrane of the retina. The external limiting membrane is the zone of adhesion (tight junctions) between photoreceptors and Müller cells. Müller cells are functionally similar to neuroglia in that they support, nourish, and insulate the neurons and fibers of the retina.
Retinal diseases
In the structure of ophthalmological diseases and pathologies, the incidence of retinal diseases, according to rough estimates, is not ˃1%. The most common violations can be divided into several groups:
- dystrophic pathologies of the retina (congenital or acquired);
- inflammatory diseases;
- damage due to eye injuries;
- anomalies associated with concomitant diseases - the cardiovascular system, endocrine disorders, pathological neoplasms, etc.
General symptoms
With abnormal functioning of the retina, patients report similar symptoms:
- visual acuity decreases;
- abnormalities in the visual field appear (it narrows, “blind” areas are observed - scotomas);
- the eye's adaptation to darkness worsens;
- color vision anomalies occur.
As a rule, there is no pain in diseases of the retina - nerve impulses are not transmitted due to the lack of sensitive innervation.
Some diseases
As an example, we should consider several of the most common pathologies of the retina:
- impaired peripheral vision - retinal pigmentary degeneration, which is a hereditary disease;
- impairment of central vision - retinal macular degeneration (macula cells die or are damaged);
- anomaly of retinal photoreceptors - rod-cone dystrophy;
- retinal detachment - it separates from the back wall of the eyeball;
- malignant neoplasms – retinoblastoma (a tumor forms in the retina);
- pathology of the vascular system of the central zone of the retina - macular degeneration.
By the very principle of existence - architectonics, the retina is comparable to the brain: its blood supply is formed according to a similar pattern, the complexity of the structure and the multiplicity of structural units provide rich functionality in the process of adequate transmission and perception of visual images of the surrounding world. This is the reason for the special exclusivity of the retina in the functioning of the human visual system.
Related articles:
- Structure of the human eye: photo with description
- Eye pressure: normal, symptoms of increase, treatment
- Lens of the eye: structure, functions, replacement surgery (price, consequences)
- Optic nerve: functions, diseases, treatment
How does the retina work? Retinal histophysiology
Light passes through the layers of the retina
to the rods and cones, where it is absorbed, triggering a series of reactions that enable vision, an extremely sensitive process. Experimental data show that one photon is sufficient to generate a receptor potential in a rod cell. Light causes bleaching of visual pigments, and this photochemical process is enhanced by mechanisms that cause local production of signals that are subsequently transmitted to the brain.
Rod cell visual pigment
- rhodopsin, consists of vitamin A aldehyde (retinaldehyde) associated with specific proteins known as opsins. Because rods have lower resolution, they produce images without distinct details; they are also color insensitive. Cones, on the other hand, have a higher threshold and are responsible for sharp images and color vision. In humans, they contain three incompletely characterized pigments (iodopsins) that provide the chemical basis for the classical three-color theory of color vision.
When the light affects
on rhodopsin molecules, retinaldehyde undergoes isomerization, converting from cis to trans form. This change leads to the dissociation of retinaldehyde and opsin, a reaction called bleaching. Discoloration of the visual pigment incorporated into the membrane discs increases the conductivity of the disc membranes to calcium and causes calcium diffusion into the intracellular space of the outer segment of the photoreceptor.
Calcium affects the cell membrane
, reducing its permeability to sodium ions, and causes hyperpolarization of the cell. Electrical signals resulting from the closure of these sodium channels propagate into the inner segment and through gap junctions to neighboring cells.
At the second stage, visual pigment
is reduced and calcium ions are transported back into the discs in an energy-intensive process. High energy requirements explain the abundance of mitochondria near the light-sensitive region of rod and cone cells. Unlike what happens in other receptors, where action potentials are generated by depolarization of the cell, rod and cone cells hyperpolarize when exposed to light.
This signal is transmitted to bipolar
, amacrine and horizontal cells, and then to ganglion cells. Only ganglion cells generate action potentials that travel along their axons, which carry information to the brain.
Clinical observations of retinal damage
when it is detached, they show that light-sensitive cells receive nutrients from the choriocapillaris layer. The superficial location of the retinal vessels makes it possible to easily study them using an ophthalmoscope. Such research is of great value in the diagnosis and evaluation of diseases affecting blood vessels, such as diabetes and high blood pressure.
At the rear pole of the optical axis
The fovea is located - a shallow depression, in the center of which the retina has a very small thickness. This is because bipolar and ganglion cells accumulate along the periphery of this recess, so that the central part contains only cone cells.
Cone cells in the fovea
- long and narrow, which is why they resemble rod cells. Thanks to this adaptation, the cone cells are located more closely, and, consequently, visual acuity increases. In this area, blood vessels do not pass over the light-sensitive cells. Light falls directly on the cones in the central part of the fovea, which causes extremely high visual acuity in this part of the retina.